Son células que se encuentran
normalmente en los tejidos conectivos, derivan de células mieloides y poseen
numerosos gránulos en su interior con sustancias como la histamina y la
heparina. Los mastocitos están relacionados con el sistema inmune, más
concretamente con las reacciones de hirpersensibilización y alérgicas.
Probablemente son células filogenéticamente antiguas ya que se encuentran en
todas las especies con circulación sanguínea. Tienen un aspecto redondeado y
sus gránulos presentan metacromasia (adquieren un color diferente al del
colorante).
Origen
Al igualque los basófilos, pertenecen a una misma
familia, se derivan en la médula ósea a partir de la célula progenitora de la
línea mieloide. Su producción está controlada por la acción de IL-9, reforzada
por la IL-3, producidas por los linfocitos T. La IL-4 es un factor regulador de
la producción y maduración de los mastocitos. Los mastocitos se encuentran en
los tejidos y epitelios mucosos.
Estructura
Su citoplasma contiene numerosos
gránulos rodeados de membrana que están llenos de proteoglucanos ácidos que se
unen a pigmentos básicos. Los mastocitos expresan en la membrana receptores de
afinidad alta para un tipo de anticuerpo llamado IgE y suelen estar cubiertos por
ellos. Cuando los anticuerpos que están en la superficie del mastocito se unen
al antígeno, se inducen acontecimientos transmisores de señales que conducen a
la liberación del contenido del gránulo citoplásmico hacia el espacio
extracelular. El contenido granular liberado, incluida la histamina, promueve cambios
en los vasos sanguíneos que producen inflamación.
Funciones
Los mastocitos actúan como centinelas
en los tejidos, donde reconocen los productos microbianos y responden
produciendo citocinas y otros mediadores que inducen la inflamación. Estas células
proporcionan una defensa frente a los helmintos y otros microbios, pero son
también responsables de los síntomas de las enfermedades alérgicas.
Los mastocitos en su degranulación
liberan mediadores primarios o prefabricados como la histamina, varias
proteasas y TNFα e inician la síntesis de los llamados mediadores secundarios, tales
como prostaglandinas, leucotrienos, quimioquinas, citoquinas, y factores
estimuladores del crecimiento de los fibroblastos y de la angiogénesis (formación
de nuevos vasos).
Referencias bibliográficas:
Abbas, A. K., Lichtman, A. H., & Pillai, S. (2015). Octava edición. Inmunología
celular y molecular. Barcelona, España: Elsevier.
Rojas M., W., Anaya C.,
J. M., et al. (2010). Decimoquinta edición. Inmunología de Rojas. Medellín, Colombia:
Corporación para Investigaciones Biológicas.
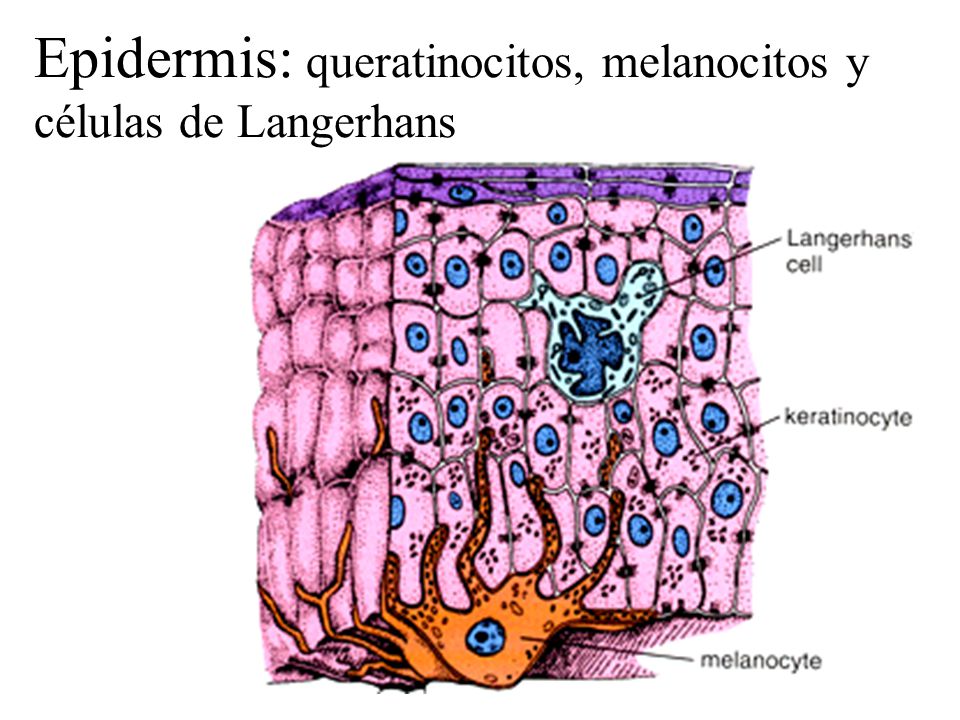
Las células dendríticas deriva del griego
δένδρον , que significa
árbol, por sus abundantes ramificaciones. También es conocida como DC (por sus
siglas en inglés), son un tipo de células especializadas características del sistema
inmunitario de mamíferos.
Aunque la célula dendrítica forme parte de la inmunidad
innata, siendo capaces de fagocitar patógenos, su función principal es la de
procesar material antigénico, convirtiendo las proteínas en péptidos que
podrán ser expresarlo en su superficie por medio del complejo mayor de histocompatibilidad (CMH) y presentarlo a las células especializadas del
sistema inmunitario adaptativo. Es por lo que se le considera un vínculo entre
el sistema inmunitario adaptativo y el innato.
Es considerada las principales células presentadoras
antigénicas, por su capacidad de capturar, procesar y presentar antígenos de
forma óptima a linfocitos T, y generar respuestas inmunes específicas
ORIGEN:
Las DCs derivan directamente de la célula madre de la médula,
salen al torrente circulatorio y van a colonizar casi todos los tejidos, colonización
que es controlada por diferentes quimioquinas y sus respectivos receptores.
ESTRUCTURA
Las células dendríticas se caracterizan por la gran densidad
de moléculas del MHC de clase II que presentan en la membrana plasmática. Las
células de Langerhans, un tipo especializado de células dendríticas, también
presentan una elevada concentración de moléculas del MHC de clase II, pero las
caracteriza la presencia de gránulo de Birbeck, unos orgánulos exclusivos de
este tipo celular con forma de barra o de raqueta de tenis cuya función aún es
discutida. Sin embargo se ha visto que están asociados a una lectina
dependiente de calcio, la langerina, y que incluso induce su formación. De ese
modo parece que la inducción de los gránulos de Birbeck sería una consecuencia
de la función de captura de antígenos de la langerina, que permitiría
conducirlo a los gránulos y activar de ese modo al procesamiento del antígeno
por la ruta no clásica. Según el microambiente en que se encuentren, las
células dendríticas expresan diversos marcadores de superficie, por ejemplo las citosinas: IL-12, IL-1 y
TNF-α.26
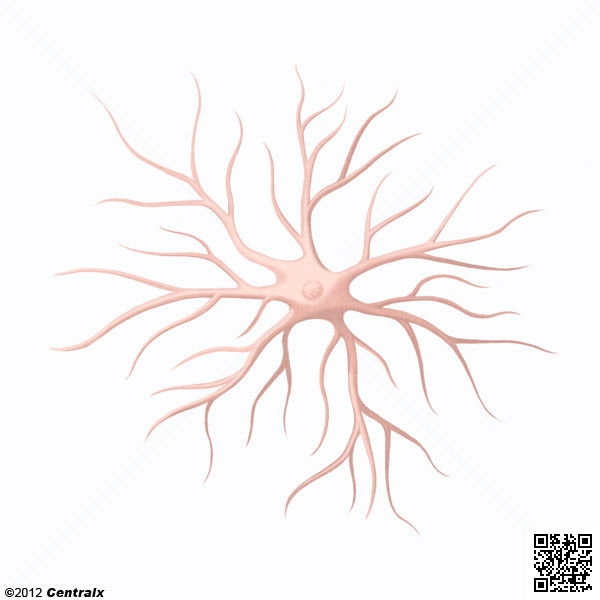
Sólo las células dendríticas inmaduras que patrullan las
regiones periféricas tienen la típica forma de estrella, que les viene dada por
unos procesos citoplasmáticos llamados dendritas, que miden más de 10 μm y que
sobresalen del cuerpo de la célula. En las células vivas, estas proyecciones
están en constante movimiento, retorciéndose, contrayéndose y volviéndose a extender
por el lado contrario. En esta fase las células tienen una gran cantidad de
vesículas endocíticas ricas en proteínas lisosomales. Este fenotipo inmaduro se
caracteriza por la escasez de proteínas del MHC y la ausencia de moléculas B7
coestimuladora.
Durante la migración de la célula dendrítica hacia órganos
linfáticos secundarios, después de haber capturado un antígeno durante una
infección, la morfología de las células dendríticas cambia. Las dendríticas son
remplazadas por pliegues y proyecciones membranales semejantes a velos, por lo
que se las puede llamar «células veliformes». Al mismo tiempo, pierden la capacidad de fagocitar y procesar
antígenos.
Finalmente, en los ganglios linfáticos u otros órganos
linfoides diana están presentes como células dendríticas maduras. Estas células
dendríticas segregan grandes cantidades de péptidos cargados de complejo MHC
II, así como varias moléculas coestimuladora, especialmente la B7. Ambas cosas
son necesarias para permitir el reconocimiento de los antígenos procesados por
parte de los linfocitos T, así como la activación de estos últimos. Mientras
que los péptidos y complejos MHC interactúan con los receptores de los
linfocitos T, las moléculas B7 proporcionan la segunda señal necesaria para
unirse a los antígenos CD28 de los linfocitos T. Las células dendríticas
maduras son capaces de estimular linfocitos T CD4 + TH1 con una gran eficacia.
SUBPOBLACIONES Y LOCALIZACIONES
Se han identificado tres subpoblaciones con fenotipos diferentes
según su origen y el tejido u órgano en donde se ubiquen.
1. DCs mieloides, mDCs. Se originan en los monoblastos o en los monocitos que
al pasar del torrente sanguíneo a los tejidos se convierten en macrófagos o en
DCs. Expresan marcadores mieloides (CD11c, CD33, CD13) y moléculas HLA-I y
HLA-II. Circulan en la sangre y en casi todos los tejidos. Tienen una gran
capacidad migratoria porque expresan Ralph Steinman, recibió
el premio Nobel en el 2011 por el descubrimiento y estudio de las DCs, lo que
indica la importancia que ellas tienen en la respuesta inmune. CCR5, CCR7,
CCR8, CCR9, que les permiten responder al llamado de las quimioquinas originadas
en diferentes tejidos. Con el CCR7, por ejemplo, responden a las CCL19 y CCL21
generadas en los ganglios linfáticos a donde viajan una vez capturan un Ag en
la periferia. Tienen distintos fenotipos y funciones según el órgano o tejido
que colonicen: en la dermis expresan varias lectinas, producen IL-6 e IL-12 y
presentan Ags a los LsB de memoria para estimularlos a producir Igs. Como
expresan CD1a (ver más adelante las moléculas CD1, sección 8-VII-B) presentan inmunógenos
lipídicos y al hacerlo inician la producción de IL-15 que activa a las demás células
del sistema inmune innato; las que van al intestino expresan CD103, que hace
parte de una integrina que facilita la ubicación interepitelial en las mucosas;
las que se ubican en las amígdalas y en el bazo expresan CD 141 e inducen la
producción local de IgA e IgM.
2. Células de Langerhans. Se originan en la etapa embrionaria y migran a la
epidermis en donde se multiplican localmente para perpetuarse sin necesidad de
que lleguen remplazos desde la médula ósea. Representan de un 5% a 8% de las
células de la epidermis. Expresan langerina (CD207), que es una lectina que permite
el reconocimiento de Ags glucoprotéicos presentes en algunos virus. La caderina
E las mantiene unidas a los queratinocitos. Son migratorias y llevan los Ags de
la periferia a los ganglios linfáticos para inducir la iniciación de una
respuesta inmune específica. No viajan al bazo. Para varios autores estas
células pertenecen más a la línea de Møs tisulares que a DCs.
3. DCs plasmocitoides, pDCs. Se originan en la médula ósea de la
línea linfoide. Morfológicamente recuerdan a las células plasmáticas. Están
ampliamente distribuidas en el organismo y ante la presencia de un virus, capturan
sus ácidos nucleicos por medio de TLR7 y TLR9, producen grandes cantidades de
IFNs tipo I, hasta 1000 veces más que otras células del sistema inmune.
Participan activamente en la patogénesis de todas las enfermedades autoinmunes.
4. Astrocitos. Son las células presentadoras de Ags el sistema
nervioso central y posiblemente pertenecen al sistema de las DCs.
moléculas receptoras que se expresan en la célula dendritica y función.
Los monocitos son un tipo de glóbulos blancos agranulocitos, Y se consideran los leucocitos de mayor tamaño, llegando a medir 10 a 15 um
ORIGEN
Se originan en la médula ósea a partir de la célula madre o pluripotencial por efecto de las citoquinas GM-CSF, M-CSF e IL-3, secretadas por diversas células, pero especialmente por los LsT. Una vez que se han multiplicado en la medula ósea, salen a la circulación para pasar luego a los tejidos.
Cuando los Monocitos migran de la sangre a los tejidos se transforman en Macrófagos o en células dendríticas, células que se distinguen por la expresión de determinados marcadores de superficie o CDs. Los Macrófagos, tienen un mayor tamaño que los Monocitos, 15-80 μm, forman poblaciones celulares heterogéneas, distribuidas en diferentes tejidos y órganos responsables de procesos de defensa inmunológica.
Los monocitos forman dos subpoblaciones distinguibles por marcadores de la superficie celular y funciones.
1.integrada por los que expresan la molécula CX3CR-1, la cual permite unirse a otras que los atraen al endotelio vascular para facilitar su paso del torrente circulatorio a los tejidos a cumplir su función de patrullaje en búsqueda de algo anormal.
2.integrada por los que expresan la molécula CCR2 y se conocen como monocitos inflamatorios, por cuanto únicamente pasan a los tejidos cuando son "llamados" por moléculas quimiotácticas que inducen en las células del tejido vascular incremento en la expresión de moléculas
adherencia para facilitar el paso masivo de este grupo de monocitos.
ESTRUCTURA
Durante el proceso de maduración los Monocitos acumulan un contenido importante de microfibrillas y microtúbulos en su citoplasma que les proporcionan una gran movilidad y capacidad de fagocitosis. En sus sacos lisosomales almacenan una gran cantidad de gránulos que contienen un arsenal enzimático para destruir todo lo extraño consistente en: lisozima, proteasas neutras, hidrolasas ácidas y arginasa, enzimas que destruyen componentes celulares y tisulares y coadyuvan a la generación de metabolitos activos del oxígeno y del nitrógeno. Este arsenal de “armas de destrucción” es empleado cuando el Monocito se trasforma en Macrófago.
FUNCIÓN
Como sabemos el monocito puede diferenciarse debido a distintos estímulos, tanto en macrófago como en célula dendrítica. Por lo que se puede decir que el monocito desempeña la función de fagocitosis, si el monocito se diferenció en macrófago, o como una célula presentadora de antígenos a los linfocitos T, si se diferenció en célula dendrítica.
Los neutrófilos son leucocitos de tipo granulocito, también llamados leucocitos polimorfonucleares, son la población más abundante de leucocitos circulantes en el ser humano.
Los neutrófilos derivan de la célula madre de la médula ósea, luego de un proceso progresivo de multiplicación y diferenciación. La médula ósea produce siete millones de neutrófilos por minuto, gran parte de los cuales se acumulan como reserva para entrar en circulación cuando un proceso infeccioso o inflamatorio lo demande.
Su producción está controlada por los siguientes factores:
1.el G-CSF o estimulador de la formación de colonias de granulocitos que se produce como respuesta a la IL-17 sintetizada por diferentes Ls;
2. la IL-3 secretada por Møs y Ls y que actúa sobre la célula madre de la médula;
3. la CXCL12 producida por el estroma de la médula que retiene a los neutrofilos en este órgano a fin de asegurar una buena reserva de los mismos;
4. un factor de liberación producido por los Mons que facilita su salida de la médula ósea.
ESTRUCTURA
Los neutrófilos son células esféricas, de 12 a 15 μm de díametro, tienen un núcleo segmentado en tres a cinco lóbulos conectados, de aquí el sinónimo leucocito polimorfonuclear y tiene un citoplasma rico en gránulos, que se pueden clasificar en dos tipos:
•La gran mayoría son gránulos específicos (son los que están en mayor cantidad), están llenos de enzimas como la lisozima, la colagenasa y la elastasa. Estos gránulos no se tiñen intensamente con las tinciones básicas ni ácidas (hematoxilina y eosina, respectivamente), lo que distingue a los gránulos del neutrófilo de los de otros tipos de granulocitos circulantes, llamados basófilos y eosinófilos.
•El resto de los gránulos de los neutrófilos, llamados gránulos azurófilos, son lisosomas que contienen enzimas y otras sustancias microbicidas, como las defensinas y las catelicidinas.
posen un citoesqueleto muy desarrollado que les permite cumplir las funciones de migración a los tejidos, patrullaje, fagocitosis y desgranulación.
FUNCIÓNES
-Componentes esenciales del Sistema Inmune Natural. Son las principales células fagocíticas encontradas en sangre periférica; correspondiéndose con un 50-70% del total de células de la serie blanca. Se les considera la primera línea de defensa contra infecciones bacterianas y fúngicas (además de las barreras naturales).
-Participan en el proceso de inflamación cuando algún tejido está dañado o infectado estos migran desde la circulación hacia los tejidos, donde reconocen moléculas especificas en la superficie del antígeno o tejido dañado e inician el proceso de fagocitosis (proceso facilitado por proteínas opsónicas), donde su capacidad microbicida permite la destrucción de una gran cantidad de microorganismos en segundos, por la acción confluente de mecanismos citotóxicos dependientes e independientes de reactivos del oxígeno
-Tienen en su membrana lípidos que son fuente importante de mediadores de la inflamación. De ellos se origina parte de los leucotrienos y prostaglandinas, asi como el factor activador de las plaquetas (PAF), que participa en varios de los mecanismos de defensa y en la inmunopatología de afecciones autoinmunes.
El neutrófilo maduro es una célula terminal, que muere por lisis una vez cumple su función o por apoptosis si pasados siete días no ha encontrado qué fagocitar.
RECEPTORES
Una vez que se llega a zona objetivo (zona lesionado o infectada), los neutrófilos deben reconocer de forma específica el agente ofensivo, antes de proceder a eliminarlo. Los neutrófilos (así como otras células) presentan en su superficie externa receptores de membrana que les permite reconocer el agente externo y activar los procesos de fagocitosis. Los tipos de receptores más importantes son:
Receptores para componentes microbianos
Receptores de tipo Toll (TLR): reconocen componentes de diferentes tipos de microbios: lipopolisacáridos bacterianos, proteoglicanos bacterianos, nucleótidos CpG no metilados (frecuentes en bacterias) o ARN de doble hebra (producido por algunos virus). Los TLR están presentes en la superficie celular, y en los endosomas, por lo que pueden detectar microbios extracelulares y fagocitados. Estos receptores activan kinasas que estimulan la producción de sustancias microbicidas.
Receptores acoplados a proteínas G
Algunos de estos receptores reconocen péptidos que contengan fragmentos de N-formilmetionina. Otros receptores reconocen quimioquinas, fragmentos del sistema del complemento, como C5a, y mediadores lipídicos, como PAF, prostaglandinas o leucotrienos, todos los cuales se producen en el contexto de daño celular. Unión del ligando a estos receptores induce la migración y la producción de sustancias microbicidas.
Receptores para opsoninas
Los leucocitos expresan receptores para opsoninas, proteínas de defensa que recubren los microbios mediante el proceso de opsonización. Estas sustancias incluyen anticuerpos, proteínas del sistema del complemento y lectinas. Una de las formas más eficaces de mejorar la fagocitosis de una partícula es recubrirla con anticuerpos tipo IgG específicas para esa partícula. Los IgG son reconocidos por los receptores de alta afinidad para Fcγ de los fagocitos, denominados FcγR. Asimismo, C3b es también una opsonina, y los fagocitos expresan un receptor, CR1, capaz de detectarlo. La unión de las opsoninas a sus receptores en los fagocitos promueven la fagocitosis y activan los leucocitos.
Receptores para citoquinas
Estos receptores sus estimulados por citoquinas producidas en presencia de microbios. La más importante de estas citoquinas es el interferón-γ (IFN-γ), segregado por las células NK activadas por microbios y por linfocitos T activados por antígenos durante la respuesta inmune adaptativa. El IFN-γ es el principal agente activador de los macrófagos.
en esta imagen podemos ver algunos de los receptores y la función que estos tienen en el neutrófilo. podemos ver como se representa la secreción de citocinas y quimiocinas asi como las funciones del neutrofilo por medio de una flecha que sale del neutrofilo.
Los linfocitos NK (Natural Killer), son un componente central de la respuesta inmune innata y constituyen la primera barrera de defensa ante una gran variedad de tumores y agentes patógenos microbianos. Su rol, es análogo al de los linfocitos T citotóxicos en la respuesta inmune adaptativa.
En el ser humano, los linfocitos NK representan del 5 al 15 porciento de la concentración total de linfocitos sanguíneos. La mayoría de los linfocitos NK, se presentan con relativa abundacia en la Médula ósea, hígado, útero, bazo, y pulmones, así como una menor cantidad presente en tejido linfoide secundario y el timo.
La descripción clásica de los linfocitos NK se basa en la expresión de CD16 (Potenciador de fagocitocis y citotoxicidad mediante la unión de IgG), y CD56 (citotoxicidad, adhesión celular y hematopoyésis), existiendo tres fenotipos celulares los cuales son los que se muestran en la siguiente tabla.
Estructura:
Receptores de células NK
Los receptores de los linfocitos NK se pueden clasificar en base a su función. Mientras que otras células citotóxicas llevan a cabo su función a través de ligandos que directamente indican que una célula está infectada o a través de la expresión de MHC, los linfocitos NK llevan a cabo su función basándose en la estimulación de sus receptores, inhibidores y activadores.
Por ejemplo, si el receptor inhibidor recibe una señal más prominente, la actividad del linfocito NK será inhibida, y si sus receptores activadores reciben una mayor estimulación, entonces su actividad se llevará a cabo.
Origen:
Los linfocitos NK constituyen el tercer tipo de células en que el progenitor linfoide común se diferencía (siendo los tros 2 linfocito T y B) en el proceso de Linfopoyesis,en la Hematopoyesis. Maduran y se diferencían en la médula ósea, nódulos linfáticos, bazo, amígdalas, y en el timo, en donde después entran a la circulación.
Se ha descubierto reciéntemente, que células precursoras hematopoyéticas que expresan CD34 se diferencían en células NK pasan a expresar CD56 dentro de los nódulos linfáticos. Un proceso similar ocurre en el hígado. Sin embargo, la médula ósea sigue siendo el sitio predilecto para el desarrollo, maduración y diferenciación de los linfocitos NK, debido en parte también, a las constantes señales que su precursor recibe por parte de las células estromales y hematopoyéticas, presentes en mayor medida en la médula ósea.
La diferencia entre los linfocitos NK y TNK consiste en que, los linfocitos TNK poseen receptores de antigenos de células T (TCR) y CD3 (Transducción de señales, interacción antigénica) mientras que los linfocitos NK no.
Desarrollos De células T y Células NK
a) En la médula ósea, las células NK en desarrollo interactúan con el MHC clase I en células estromales y hematopoyéticas a través de receptores inhibidores, y el número de interacciones determina el grado de respuesta
b) El desarrollo de células NK que interactúan con ligandos propios o virales a través de receptores activadores se volverán anérgicas o hiporreactivas
c) En el timo, las células T en desarrollo interactúan con el péptido propio / MHC en células epiteliales y hematopoyéticas a través de receptores de células T, y la afinidad o avidez de estas interacciones determina la supervivencia y la exportación a la periferia
Funciones:
Sus características funcionales, incluyen la producción y secreción de IFNy y TNFa, así como G-CSF. Podrían también, producir cierta variedad de quimocinas, entre ellas IL-8, y en condiciones muy especializadas y limitadas IL-1, IL-10 e IL-6.
Los linfocitos NK proveen una respuesta rápida ante células infectadas por virus, actuando alrededor de 3 días después de la infección.
Comúnmente, las células del sistema inmune detectan el complejo mayor de histocompatibilidad (MHC) que se presenta en la superficie de la membrana de las células infectadas. Sin embargo, los linfocitos NK son únicos, ya que pueden reconocer células estresadas o infectados incluso en la ausencia de anticuerpos y MHC, permitiendo una respuesta mucho más rápida.
Apoptosis celular mediada por gránulos
Los linfocitos NK son citotóxicos; pequeños gránulos en su citoplasma contienen proteínas como perforinas y proteasas. Tras su secreción en proximidad a una célula diana, las perforinas crean poros en la membrana de esta, creando canales acuosos por los cuales el agua y otras granzimas podrán entrar, causando ya sea apoptosis o lisis celular osmótica.
Citotoxicidad celular dependiente de anticuerpos
Las celulas infectadas son opsonizadas por anticuerpos. Los anticuerpos unidos a células infectadas, son reconocidos por el CD16 en la membrana de los linfocitos NK, resultando en su activación, secreción de granulos citolíticos y posterior apoptosis celular.
Activación inducida por citocinas
Las citocinas juegan un papel importante en la activación del linfocito NK, ya que son moleculas que ayudan a señalar la presencia de antigenos en un área afectada. Las interleucinas presentes en la activación del linfocito NK incluyen IL-12, IL-15, IL-18, IL-2 y CCL5. Se activan en respuesta a interferones o a citocinas derivadas del macrófago.
Vigilancia tumoral
Los linfocitos NK carecen de receptores antígeno-específicos, por lo que reaccionan ante el agente extraño inmediatamente.Tienen un rol importante en la inmunovigilancia tumoral induciendo directamente la muerte de células tumorales, incluso en la ausencia de moléculas de adhesión y péptidos antigénicos,esta acción de los linfocitos NK es crítica porque los linfocitos T, en ausencia de antígenos de superficie, no son capaces de reconocer agentes patógenos.
Asímismo, si una célula tumoral no produce un proceso inflamatorio, será tratada como células propia y no habrá actividad alguna de células T. Para que las células T puedan activarse, es necesaria la secreción de IFN, TNF e IL-10 por parte de los linfocitos NK, que mediante las citocinas mencionadas, activará a los macrófagos, células dendríticas y neutrófilos, lo cual habilitará respuestos antígeno-específicas por parte de los linfocitos T y B.
Son polimorfonucleares de 12 micras de diámetro y su núcleo tiene una forma que recuerda a una S, es lobulado y se divide de 1 a 3 lóbulos., se caracterizan por presentar en su citoplasma numerosos gránulos redondeados, unos mil por célula, que miden en promedio media micra y que con los colorantes de anilinas básicas se tiñen metacromáticamente de púrpura.
Esta coloración se debe a la presencia de heparina que constituye el 30% del material que integra los gránulos. El resto está formado por histamina y enzimas como histidina descarboxilasa, proteasas, fosfatidasa A, fosfatasa ácida y alcalina, beta glucorunidasa, dopa descarboxilasa, citocromo-oxidasa, descarboxilasa de aminoácidos y factor activador de las plaquetas.
Tienen una activa participación en la respuesta inmunitaria, a través de la liberación de histamina, serotonina en bajas concentraciones, y otras sustancias químicas, además tiene gránulos de dos clases:
Gránulos azurófilos: Contienen lisosomas, que a su vez estos contienen hidrolasas ácidas.
Gránulos específicos o secundarios: contienen histamina (vasodilatador), heparán sulfato (vasodilatador), heparina (anticoagulante) y leucotrienos (hacen contraer el músculo liso de las vías aéreas).
Origen:
Son leucocitos circulantes derivados de médula ósea. Son células mononucleares con un alto contenido granular. Su diferenciación a partir de progenitores mieloides está mediada por IL-3. Ésta actúa sobre la cadena α del receptor para IL-3 (también conocido como CD123), lo que permite distinguirlos de los mastocitos.
Funciones:
Los basófilos además de poseer gránulos en su interior poseen receptores de IgE (inmunoglobulina E), aquella inmunoglobulina relacionada con las alergias. Por eso el basófilo participa en la respuesta inflamatoria.
Los basófilos constituyen menos del 1% de los leucocitos en humanos, pero son las únicas células circulantes que contienen histamina, estos comparten muchas similitudes con los mastocitos residentes en los tejidos, activándose cuando se produce entrecruzamiento del antígeno con el receptor FcεRI de IgE.
El entrecruzamiento provoca la rápida degranulación y liberación del contenido intracelular. Los basófilos también pueden activarse sin la presencia de IgE, gracias a mediadores inflamatorios como los factores del complemento (C5a y C3a), MBP, PAF y quimiocinas.
Los basófilos son reclutados a los sitios de inflamación donde pueden ser activados directamente por una variedad de patrones moleculares asociados a patógenos (o PAMPs, por sus siglas en inglés), así como por entrecruzamiento de IgE.
Cuando son estimulados, liberan su contenido granular conteniendo histamina y producen LTC4 (leucotrieno 4). Además, también son capaces de producir citocinas, principalmente IL-4 e IL-13, así como MIP-1α. Por lo tanto, estas células ayudan al sistema inmunitario a través de la liberación de distintos componentes tales como citocinas, leucotrienos e histamina. En particular, los basófilos son realmente importantes en respuestas inmunitarias frente a parásitos, incluyendo garrapatas y filarias.
Los basófilos ya han sido implicados en la patogénesis de la enfermedad alérgica, ya que se han encontrado niveles altos de los mediadores que ambos producen en tejidos afectados por la alergia.
Los basófilos son la fuente principal de producción de IL-4 en las respuestas inmunitarias iniciales, una citocina que promueve la diferenciación de las células T hacia un fenotipo Th2. Además, una vez que el antígeno ha entrado en el organismo, estas células pueden rápidamente ser reclutadas a piel, pulmón o mucosa nasal, por lo que se encuentran elevados en dermatitis atópica, asma, rinitis alérgica y pólipos nasales.
Referencias:
Davoine F, Lacy P. 2014. Eosinophil cytokines, chemokines, and growth factors: emerging roles in immunity. Frontiers in immunology 10. doi:10.3389/fimmu.2014.00570.
Muniz VS, Weller PF, Neves JS. 2012. Eosinophil crystalloid granules: structure, function, and beyond. J Leukoc Biol. 92:281-288.
Padigel UM, Nolan TJ, Schad GA, Abraham D. 2006. Eosinophils can function as antigen-presenting cells to induce primary and secondary immune responses to Strongyloides stercoralis. Infection and immunity. 74(6): 3232–3238.
Rothenberg ME, Hogan SP. 2006. The eosinophil. Annual review of immunology. 6. 24:147-174.
William Rojas M., Luis Miguel Gómez O.. (2012). Inmunología de Rojas. México: CIB.